
4 Feb 2021
Post by Valentina Larcinese

Questo articolo, e la serie a cui appartiene, trae ispirazione dal documento “Natural selection, selective breeding, and the evolution of resistance of honeybees (Apis mellifera) against Varroa ” pubblicato il 18 maggio 2020. L’intento è di fornirne una versione semplice e fruibile per tutti.
La parte dedicata ai virus presenti nelle regine è tratta da due articoli pubblicati sull’ American Bee Journal (“Queen Forensics” – Luglio 2019 e “The virus hidden in our queen” – Febbraio 2021 ) di Allison McAfee, ricercatrice presso l’University of British Columbia dove studia i meccanismi del comportamento igienico delle api.
Cominciamo ad analizzare alcuni degli aspetti della resistenza e della tolleranza a Varroa:
- 1. Resistenza e tolleranza contro i virus associati alla varroa
- 2. Caratteristiche biologiche presenti nelle colonie resistenti e tolleranti a varroa
2.a Tendenza alla sciamatura
2.b Accoppiamento delle api, struttura della popolazione e selezione naturale.
Infine, facciamo una apparente digressione su un aspetto poco noto che possiamo ritenere un feedback negativo dell’interazione virus/varroa: ovvero, il fatto che le regine di apis mellifera possano essere anch’esse soggette all’aggressione di virus, che ne pregiudicano la funzionalità ed efficienza, rendendo più arduo per la colonia una spinta selettiva verso una maggiore tolleranza o resistenza a varroa.
Resistenza e tolleranza contro i virus associati alla varroa
Un motivo importante per cui le colonie infestate da varroa collassano è che la varroa attiva un’infezione da sindrome del virus delle ali deformi (DWV) e incrementa i valori di altri virus come la covata a sacco (SBV) e il virus della cella reale nera (BQCV).
Questi virus sono stati associati alla morte di milioni di colonie di api europee nel mondo. Le api, in assenza di questi virus, possono sopportare una maggiore popolazione di varroa prima di collassare.
L’interazione della varroa con le api affette da DWV in qualche modo porta i virus a essere molto più aggressivi. Questo non accade nelle popolazioni ‘chiuse’ (leggi isolate) perché la trasmissione verticale (ossia alla progenie) favorisce forme meno aggressive di DWV. I casi più noti sono le popolazioni presenti nell’isola di Ferdinando de Horona in Brasile, nell’isola di Gotland in Svezia, a Swidon in Inghilterra e nell’ Arnot Forest nello Stato di New York, USA.
Le popolazioni di Ferdinando de Horona, Swidon e Gotland sono frutto di esperimenti in cui si è verificata la capacità di alcune colonie di sopravvivere alla varroa nonostante l’assenza di trattamenti acaricidi; mentre la popolazione della Arnot Forest si compone di colonie selvatiche.
La minore virulenza del parassita favorisce l’incremento della tolleranza /resistenza delle api alla varroa. Inoltre, il VSH favorisce l’evoluzione di una più bassa virulenza del DWV e di altri virus . Le api igieniche hanno come obiettivo preferito le pupe che hanno subito danni da una forma aggressiva di virus e quindi può essere determinante nel rendere dominante (leggere “selezionare”) un tipo di virus meno virulento.
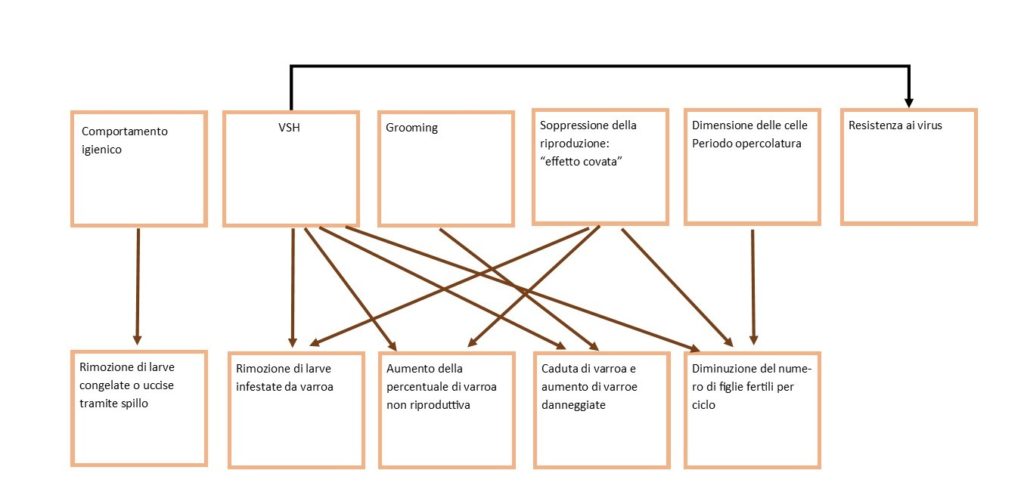
Caratteristiche biologiche che possono ostacolare la crescita della popolazione della varroa
Tendenza alla sciamatura
La grandezza di una colonia e la frequenza di sciamatura sono tratti biologici delle api che influenzano la crescita della popolazione della varroa. Sebbene questi tratti siano in qualche misura ereditabili, sono ampiamente determinati da fattori ambientali come la misura del nido e l’abbondanza di cibo.
Il meccanismo per cui le colonie più piccole e più sciamanti hanno una maggiore resistenza alla varroa comprende l’avere relativamente poca covata, soprattutto poca covata maschile, che limita la possibilità di riproduzione per gli acari. La frequente sciamatura può anche aiutare nel controllo degli acari perché con tale evento circa il 35% degli acari della colonia viene esportato, cioè va via con lo sciame. Inoltre la sciamatura priva temporaneamente gli acari della covata e si interrompe il loro ciclo di riproduzione incrementando la loro esposizione al grooming.
Seeley e suoi collaboratori hanno condotto diversi studi di osservazione e monitoraggio delle colonie selvatiche stabilitesi nell’Arnot Forest tramite i quali hanno verificato, con successo, l’ipotesi secondo cui la sopravvivenza di queste colonie selvatiche è dovuta, almeno in parte, alla loro abitudine di nidificare in piccole cavità e di sciamare frequentemente.
L’affollamento degli apiari incrementa ulteriormente la trasmissione della varroa tra le colonie: la distribuzione sparpagliata delle colonie selvatiche le rende meno esposte alla trasmissione orizzontale da parte di altre colonie tramite il fenomeno della deriva e del saccheggio.
Accoppiamento delle api, struttura della popolazione e selezione naturale
Negli insetti eusociali c’è una forte selezione per incrementare la diversità genotipica nelle operaie: sia per incontrare le domande di differenti compiti da svolgere, sia per mitigare gli effetti dei patogeni. Sebbene tutte le operaie di una colonia siano figlie di una stessa madre, la regina, le variazioni genetiche tra le operaie possono essere incrementate da due meccanismi:
- l’incremento del tasso di ricombinazione
- la poliandria.
Il tasso di ricombinazione nelle api è tra i più alti misurati nel regno animale e le api regine sono estremamente promiscue. Esiste una forte evidenza che la diversità genetica che è generata da una regina, attraverso la poliandria, dà beneficio alla colonia incrementando la produttività della forza lavoro.
Gli accoppiamenti delle regine avvengono in volo con 7-28 fuchi che possono provenire da colonie situate a 15 km di distanza. Le giovani vergini lasciano la colonia per il volo di accoppiamento. Nelle api l’accoppiamento avviene nell’area di congregazione fuchi, che sono visitate dai fuchi di molte altre colonie. Il numero di colonie dalle quali provengono i fuchi in una particolare area di congregazione dipende dalla densità delle colonie in un’area e dalla presenza o meno di barriere fisiche (corsi d’acqua e montagne). L’alta promiscuità delle regine, le lunghe distanze percorse dalle regine e dai fuchi e l’ampio numero di colonie dalle quali giungono i fuchi nell’area di congregazione fanno sì che le api abbiano una popolazione panmitica (avente origine da accoppiamenti casuali e liberi). Questa modalità di accoppiamento è importante per come la selezione naturale o l’allevamento selettivo procedano per la resistenza alla varroa quando alle regine è permesso l’accoppiamento libero. Mentre l’allevamento selettivo e la selezione naturale favoriscono i tratti della resistenza, l’accoppiamento delle regine con fuchi provenienti da popolazioni circostanti contrasta gli effetti della selezione locale.
Digressione: I Virus nascosti nelle regine
Non si pensa mai al fatto che le regine possano essere suscettibili alle malattie. Ai tempi d’oro le regine sopravvivevano almeno per un paio di anni, ma attualmente il 50% delle regine viene sostituito nell’arco di sei mesi. Capire quale sia la causa di questa inefficienza delle regine è davvero difficile.
Le cause possibili potrebbero essere rintracciate in:
- Pesticidi introdotti nell’alveare attraverso le bottinatrici
- Esposizione a basse temperature nei pacchi d’ape fatti a inizio primavera
- Esposizione a temperature troppo alte durante i viaggi per la pratica del nomadismo
- Trattamenti acaricidi
- Nosema
- Esaurimento dello sperma
Solo le ultime due cause possono essere indagate tramite dissezione della regina morta e osservazione al microscopio. Le altre cause non sono rilevabili al microscopio ma non significa che non lascino altre tracce più indaginose da rintracciare.
Allison McAfee ha condotto nel 2019 uno studio con lo scopo di trovare dei segnali molecolari che indichino se la regina abbia subito eventi di stress nel passato e, nel caso, capire quali. Il presupposto fondante della ricerca è che le cellule degli esseri viventi, comprese le nostre, possono rispondere a stress fisiologici attivando alcuni geni per mitigarne gli effetti (marcatori molecolari).
Temperature basse ed esposizione ai pesticidi possono causare problemi fisiologici che attivano una risposta cellulare di questo tipo.
Le regine sono altamente vulnerabili agli sbalzi di temperatura e questo può verificarsi quando sono nelle gabbiette con poche operaie di supporto nella termoregolazione. Il caldo (ossia una temperatura compresa tra 40 e 42 °C) può creare nella regina uno stress ossidativo che può causare il deperimento dello sperma nella spermateca. In assenza di un numero sufficiente di operarie per tenerla alla giusta temperatura, l’esposizione per due ore a 40 °C è sufficiente per ridurre la vitalità dello sperma di due terzi. Questo non significa che un giorno o anche tre giorni di temperature a 40°C danneggeranno la regina: le operaie impegnate nella raccolta di acqua e nella ventilazione saranno in grado di regolare la temperatura dell’alveare e salvaguardare la regina.
Le regine poco performanti, e soggette quindi a precoce sostituzione, hanno sperma con bassa vitalità e alti livelli dei marcatori molecolari per stress dovuto a shock termico. All’inizio del 2020 Alex McMenamin, impegnato nel conseguimento del dottorato alla Montana State University, ha pubblicato una ricerca mostrando che le infezioni virali e lo shock termico producono risposte simili. Se esposte alle alte temperature le api producono delle proteine che aiutano a combattere l’infezione virale.
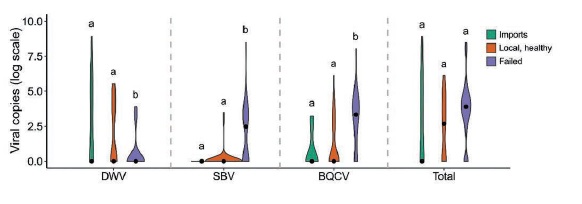
Le regine sono suscettibili alle infezioni virali – tra i più noti il virus delle ali deformi (DWV), il virus della covata a sacco (SBV) e il virus della cella reale nera (BQCV). Le regine che non muoiono a causa di una iniziale infezione da BQCV raramente mostrano segni della malattia e non mostrano differenze visibili rispetto alle altre. Le regine che sono inefficienti hanno livelli alti di SBV e BQCV ma non di DWV e la loro carica virale totale tende ad essere più alta, come si evidenzia nel grafico di seguito riportato.
Resistenza alla Varroa: quanto ne sappiamo?
Parte 1: Introduzione
Parte 2: Comportamento igienico
Parte 3: Grooming
Parte 4: Varroa non riproduttiva
Parte 5: Grandezza delle celle


0 Comments